Click on figure to open a larger version.
SUPERSEDURE AND COMPETITION
(Plate 3, Figure 11)
In the specific accounts which follow I frequently mention that a wasp or bee was superseded by another wasp or bee. In the sense used here, the phenomenon of supersedure is interpreted as being the act of taking over by a second individual of the same or of a different species a boring partially stored by a first individual. This meaning does not imply that there was necessarily any competition in this succession. It sometimes happens that a female abandons a nest before completing it - she may become the prey of a larger predator or of an internal parasite, or she may simply die of old age. I have found a few nests with the mother dead inside. In one megachilid mother there was a conopid puparium filling the abdomen; in the other nests the mother may have died from old age. The remainder of the borings in these nests may have been used subsequently by another wasp or bee days or weeks after the first individual had used them.
Sometimes there was definite competition in this supersedure (fig. 11), as in a trap from Derby in which a trypoxylonine wasp placed a paralyzed spider at the inner end of the boring: Ancistrocerus c. catskill stored cell 1, Trypargilum clavatum stored cells 2-5, and Trypoxylon frigidum stored cells 6-7. I have also observed two females of Osmia lignaria both entering the same boring with pollen and nectar loads. Also there have been some evidences of competition between two different species in a single boring. I can recall one composite nest from Florida which contained a single completed cell of Euodynerus foraminatus apopkensis at the inner end, then a partially stored cell of a species of Chalicodoma (Chelostomoides) [identifiable because inner cell end and walls were coated with resin], and then several cells of Stenodynerus saecularis rufulus. Quite possibly taking over by Stenodynerus was the result of actual competition, because the bee cell was not completely stored and it had been sealed over by the wasp.
Competition between individuals for nesting sites is a problem that could be investigated in detail in these borings by marking nesting females and observing the day-to-day activities at a station with a number of borings. I have not had time for this kind of field endeavor, but it would certainly merit investigation by someone with the requisite time, patience, and inclination.
Actually, there are many fascinating facets of behavior which could be investigated by prolonged observation at these stations. One could determine the provisioning rates for prey, the number of pollen loads to store a bee cell, the number of visits with sealing materials to make a cell partition, and myriads of others.
Two species, Solierella affinis blaisdelli and Tracheliodes amu (fig. 68), made nests without any division into cells. Several other species, Megachile (Sayapis) policaris (figs, 92-97), Isodontia (Murrayella) auripes (figs. 60, 61), and, occasionally, I. (M.) mexicana, made a large brood cell in which several larvae developed amicably without cannibalism. However, most of the wasps and bees made a series of linear cells in these borings, each cell being capped by a partition to separate it from the next cell in the series.
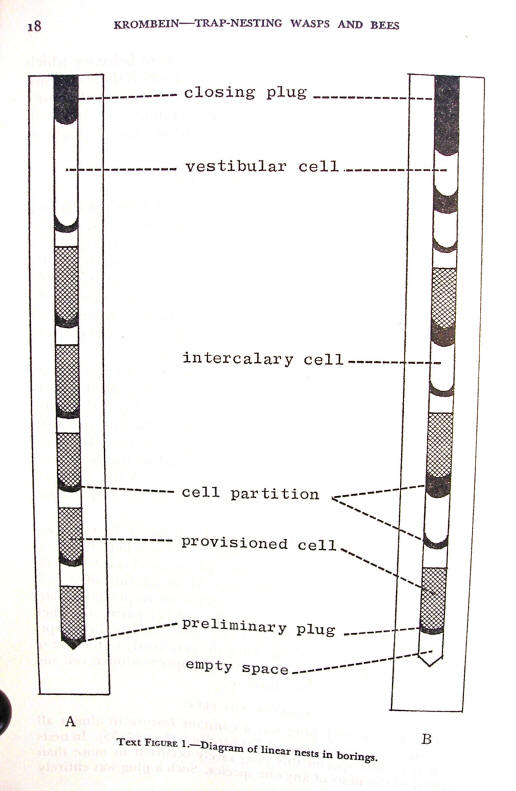
Text figure 1 illustrates two common types of serial nests. In nest A the wasp makes a preliminary plug of a small amount of mud or agglutinated sand at the inner end of the boring. Then she constructs a series of provisioned cells separated from each other by a narrow partition of the same material as the preliminary plug. Finally, at the outer end of the boring she leaves an empty space, the vestibular cell, which she seals with a thicker closing plug of the same material as the partitions between the provisioned cells.
|
|
Text Figure 1:
Diagram of linear nests in boring. Click on figure to open a larger version. |
Nest B is similar in most details. In this nest the wasp puts the preliminary plug some distance from the inner end of the boring and then constructs the first provisioned cell which she seals with the usual narrow partition. However, instead of making another provisioned cell, she leaves an empty intercalary cell which she seals with a somewhat thicker partition. Then she makes a second provisioned cell, and then another empty intercalary cell, and so forth, until she gets nearly to the end of the boring, where she makes the usual empty vestibular cell and thicker plug at the boring entrance.
PRELIMINARY PLUG
This preliminary plug was a constant feature in almost all nests of the several species of Trypargilum (figs. 53, 55). In nests of many other species this plug rarely occurred in more than a third of the nests of any one species. Such a plug was entirely lacking in nests of a few wasps. The sporadic occurrence of the plug in nests of most species suggests that it might be associated with the conformation of the inner end of the boring. Perhaps the plug was constructed to provide a smooth posterior end for the innermost cell in those borings where the inner end was roughened from protruding fibers. (See discussion below on the functions of the cell partitions.)
There was no apparent reason why this plug was made at the inner end of the boring in many nests (fig. 55) and at variable distances from the inner end in other nests of the same species (fig. 53). Individual idiosyncrasies of the nesting wasps and bees may have been the determining factor in the positioning of this partition some distance from the inner end, or the nesting females may have detected some adverse condition in the boring walls which caused them to set the preliminary plug some distance from the end.
CELL PARTITION
After the wasp or bee places a store of food in the first cell and lays an egg, or, lays an egg first and places a store of food in the cell as does the vespid wasp, she then constructs a partition to seal the cell and its contents. This partition is usually constructed several millimeters beyond the store of food. The material used in the partition varies with the individual species. Many of them use pellets of damp mud or of damp sand agglutinated by salivary secretions(?). These pellets are consolidated to form a lenticular partition. Other wasps and bees use resin, gum, leaf pulp, wood raspings from the boring walls, or even thick, compacted accumulations of various kinds of debris such as rotten wood, caterpillar droppings, small pebbles, spider webs, and earth.
Despite the disparity of materials used in their construction, these cell partitions exhibit two common features. The inner surface facing the stored cell is roughened and convex, whereas the outer surface is smooth and concave because the wasp can compact and form the material with her head. These features mean that each cell has a cylindrical form with a smooth, concave posterior wall toward the inner end of the boring and a roughened, convex anterior wall toward the nest entrance.
The cell partitions serve several obvious purposes. They offer the occupant a degree of protection from parasites and predators. They insure each larva an adequate amount of food. They prevent cannibalism between larvae in adjacent cells. An unexpected but very vital function was established by Cooper (1957) in a set of ingeniously devised experiments to determine what factors caused the larva to orient with its head toward the nest entrance, thus insuring the successful emergence of the adult wasp. Working principally with the vespid Euodynerus f. foraminatus, he was able to show that the larva oriented with its head toward roughness and convexity (i.e., the anterior end of the cell) and away from smoothness and concavity (the posterior end of the cell) so that the adult developed with its head toward the boring entrance and eventually could escape from the nest. Most mother wasps select nesting sites with a boring diameter snugly adjusted to their bodies. They cannot turn around inside the boring but must back in or back out. Consequently, their progeny must orient properly or they are doomed to death because they cannot chew through the inner blind end of the boring.
However, I found that mature larvae of two species usually or always oriented with their heads toward the blind inner end of the boring. Both species nested in 12.7-mm. borings and neither spun a cocoon. One was the large vespid wasp Monobia quadridens (Linnaeus) and the other the carpenter bee Xylocopa virginica (Linnaeus). Despite misorientation as larvae, the adults were able to orient themselves correctly. They always emerged successfully from the nests by chewing through the partitions between them and the nest entrance. Furthermore, 1 never observed a nest where the adult had first attempted to emerge toward the blind inner end before turning around and emerging toward the entrance.
Occasionally I found otherwise empty borings with just a closing partition at the entrance. For most of them it was impossible to determine the cause of this anomalous behavior. However, at one station I found four empty borings each plugged at the entrance by a spider wasp, Dipogon s. sayi. A possible explanation was disclosed in a fifth boring at this station. This boring contained two completed Dipogon cells at the inner end and then three vespid cells, the outermost still being stored with caterpillars. Apparently the supersedure by the vespid may have triggered the Dipogon female into this series of untimely and inappropriate nest closures. At times I have wondered whether some of this plugging of empty borings might not be analogous to the behavior of wrens in filling up all kinds of cavities with leaves or twigs to deny nesting sites to other birds, although I think that this explanation is rather implausible.
VESTIBULAR CELL
It is preferable to discuss the vestibular cell before the empty intercalary cells, because the vestibular type is a feature of nearly all nests whereas the intercalary is not, and because of the probable origin of the intercalary.
The vestibular cell is an empty space of varying length at or near the outer end of the boring (fig. 22). It is capped by a closing plug usually constructed of the same material as the cell partitions, although almost always twice or more as thick. Occasionally, the vestibular cell may be separated into two or more sections by narrow transverse partitions (fig. 23). It has been theorized that this cell functions to discourage penetration to the stored cells by parasites or predators. If it served this useful function at one period of nest evolution, it has now largely outlived such a purpose, because the parasites have timed their attacks to coincide with provisioning of the individual cells or they are persistent and resourceful enough to penetrate several partitions and empty spaces to get at a provisioned cell.
EMPTY INTERCALARY CELLS
In the nests of almost any species there may be an occasional empty intercalary cell between two stored cells. They occur sporadically and rarely in most species. For this reason they are of no significance in the nest architecture of most wasps, but they are probably the result of a behavioral response of the mother wasp to some factor such as the invasion of the preceding stored cell by a parasite, the discovery of mold spores or other contaminants on the boring walls in the section where the intercalary cell is made, and so forth. Conceivably, the occasional intercalary cell may actually be a vestibular cell, and may mark the spot where a second wasp superseded the maker of the first series of cells.
However, in certain wasps, particularly the vespids Monobia quadridens (fig. 29), Pachodynerus erynnis and P. astraeus, Symmorphus c. cristatus (fig. 21), and several species of Euodynerus (fig. 40), these cells were present in half or more of the nests. In some nests an intercalary cell was present between each pair of stored cells. However, these nests were in a minority, and most nests with intercalary cells had them between only some of the stored cells in a particular nest.
A peculiar but constant feature of these intercalary cells was that they were sealed by a thicker partition than the one sealing the stored cells. In other words, considered as a unit, the empty intercalary cell was indistinguishable architecturally (except for its position in a series of cells) from a vestibular cell at the nest entrance. I believe that the function of these intercalary cells has never been logically explained. Malyshev (1911, pp. 55-56) and Cooper (1953, p. 20) treated them as empty cells no different from the occasional empty cells at the inner end of the boring. I do not agree with this homology.
As I explained earlier, I consider that the plug at the inner end of the boring, whether it caps an empty space or not, serves as part of the orienting mechanism of the wasp larva in the first cell. I believe the probable explanation for the intercalary cell is that it is a relict of those days when the ancestors of these wasps may have made smaller nests consisting of just a single stored cell and a vestibular cell. During the evolution of nest building the wasps, when they found a cavity of sufficient length, put several of these single-celled nests together to form a linear series of stored cells with interspersed empty cells. As evolution progressed, more efficient use of the available space was made by the elimination of these intercalary cells, so now we find only some individuals of a few species of vespid wasps still clinging to the old habit of following each stored cell with an empty one.
Posted August 6, 2009